The billions of neurons in the human brain communicate with each other to perform computations that help us perceive and interact with the world. Neural diseases such as Parkinson’s and Alzheimer’s arise from degeneration or dysfunction of regular neural activity. To better understand what causes the breakdown of regular neural activity, researchers need to be able to record from large populations of different types of neurons at the same time.
In our lab, we have developed several genetic tools that help us address these limitations. The two main classes of tools we are developing are: (1) genetically encoded voltage indicators, which flash a certain color when a neuron is active, and (2) optogenetic actuators, which are ion pumps or channels that make a neuron active when you shine a certain color of light.
The voltage sensors we developed allow us to record neural activity with high enough temporal resolution to resolve individual action potentials and high enough sensitivity to detect subthreshold changes in a neuron’s voltage. We have developed multicolored versions of this sensor, which allow us to independently record different neural populations at the same time. Furthermore, we can combine this spectral diversity with genetic tools that allow us to target expression to genetically defined subtypes of neurons. This genetic targeting and orthogonal recording strategy enable studies which probe how populations of neurons communicate with each other, respond to particular stimuli, or modulate their activity in response to optogenetic stimulation of another neural ensemble.
 |
 |
|
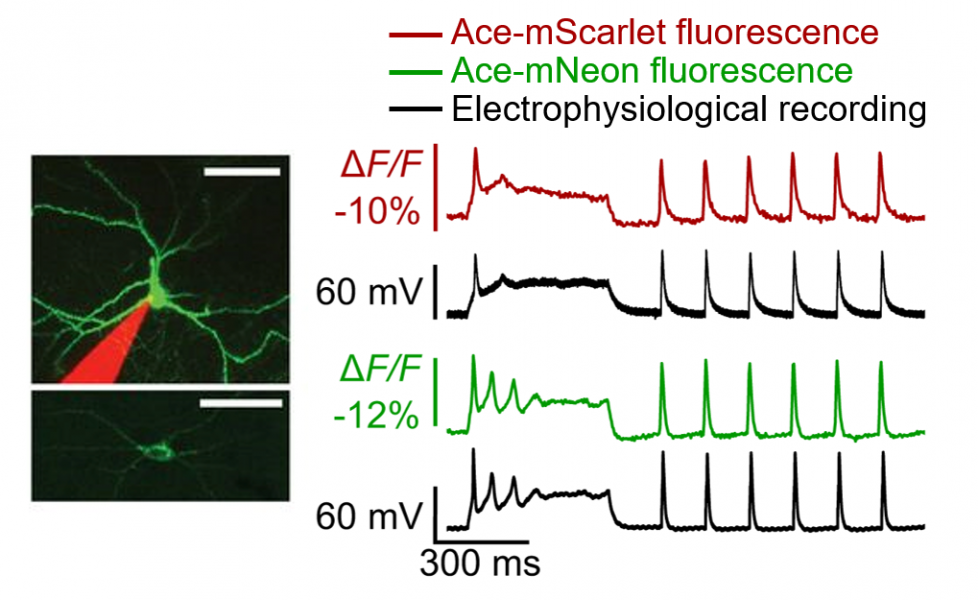
Optical recording of dendritic activity froma Purkinje neuron in a live mouse. |
Demonstration of simultaneous optical voltage recording (green/red) and electrophysiology (black) Source: Beck, C., Gong, Y. Sci Rep 9, 15878 (2019); Gong, Y. et al. Science 350, 6266 (2015) |
The optogenetic tools we developed allow us to control neural activity with high temporal precision and are sensitive enough to drive action potentials with low powered light excitation. Using rational mutagenesis of a channelrhodopsin’s important amino acids, we have developed a suite of tools with diverse kinetics and sensitivities. This diversity in the rhodopsins’ properties translates to diversity in the types of experiments for which they can be used. In general, one-photon (470 nm) stimulation requires faster ChR (toff = 2-10 ms) for high temporal resolution, and two-photon (850 nm) stimulation requires slower ChR (toff = 15-30 ms) for photocurrent accumulation.
|
|
| Left: Engineered rhodopsins expressing in a cultured neuron. Right: Different engineered rhodopsins have different off kinetics. |
